セロトニン または 5-ヒドロキシトリプタミン (5-HT) は、特に私たちの 脳 に自然に存在する 神経伝達物質 で、神経インパルスの伝達を担当します。 5-HT は 、アミノ酸 L-トリプトファンのヒドロキシル化とカルボキシル化によって生じるインドールアミンです。

 中枢神経系
(CNS)や体の他の領域(腸粘膜、
血小板
、マスト細胞などに見られるエンテロクロム親和性細胞など)におけるセロトニンの合成への最初のステップは、トリプトファンの取り込みです。 。これは、特にタンパク質食から得られるものですが、他のアミノ酸鎖に共通するキャリアによって活発に輸送されます。したがって、主に脳内のトリプトファンのレベルは、
血漿
中のトリプトファンの濃度だけでなく、同じタンパク質担体をめぐって競合する他のアミノ酸の同じ場所の濃度にも影響されます。
中枢神経系
(CNS)や体の他の領域(腸粘膜、
血小板
、マスト細胞などに見られるエンテロクロム親和性細胞など)におけるセロトニンの合成への最初のステップは、トリプトファンの取り込みです。 。これは、特にタンパク質食から得られるものですが、他のアミノ酸鎖に共通するキャリアによって活発に輸送されます。したがって、主に脳内のトリプトファンのレベルは、
血漿
中のトリプトファンの濃度だけでなく、同じタンパク質担体をめぐって競合する他のアミノ酸の同じ場所の濃度にも影響されます。
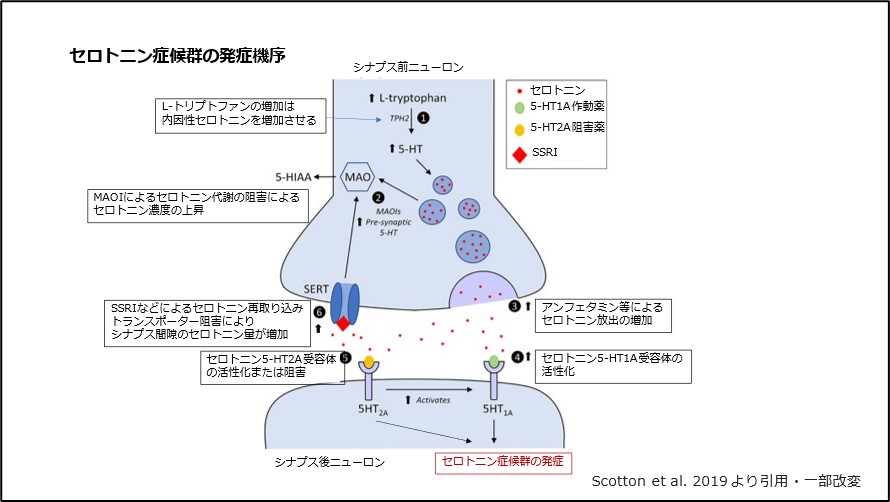
L-トリプトファンはトリプトファンヒドロキシラーゼ酵素の作用を受け、L-5ヒドロキシトリプトファンの形に変化します。その結果、後者の形態は、L-酸脱炭酸酵素の作用によってセロトニンに変換されます。この酵素は高度に分布しており、さまざまな基質に対して広範囲の特異性を持っているため、この酵素経路を通じて脳内のセロトニンレベルを制御することは事実上不可能です。
この神経伝達物質の生体内変換は肝臓またはその起源の場所で起こる可能性があり、このプロセスの主な経路は 2 段階で起こり、酵素モノアミンオキシダーゼ (MAO) が関与します。まず、MAO によってセロトニンの酸化的脱アミノ化が起こり、5-ヒドロキシインドール アセチルデヒドが生成されます。その後、酵素 アルデヒド デヒドロゲナーゼによってその主な代謝産物である 5-ヒドロキシインドール酢酸 (5-HIAA) に酸化されるか、アルデヒドレダクターゼによって 5-ヒドロキシトリプトフォール (5-HTOL) に還元されます。これらの代謝産物は変化し、尿とともに排出されることがあります。
この神経伝達物質は、さまざまな受容体との相互作用を通じてその役割を果たします。 5-HT 受容体は、その構造的および動作的特徴に基づいて 7 つの異なるクラス (5-HT 1 、5-HT 7 ) にさらに分類され、14 のサブタイプが同定されています。

5-HT 1A 受容体は、セロトニン作動性ニューロンの活性の調節を担う体性樹状突起自己受容体として機能します。どうやら、この受容体の活性化は、感情および摂食行動、認知機能、 細胞の成熟および分化を 調節するようです。 5-HT 1B および 5-HT 1D 受容体は、セロトニンおよび アセチルコリン などの他の神経伝達物質の放出を調節します。 5-HT 2 受容体は視覚野に関連しており、摂食行動を調節し、血管収縮を媒介します。一方、5-HT 3 受容体は 5-HT 放出の調節に関与しており、明らかに痛みの知覚、アセチルコリンと ドーパミン の放出のメカニズムに関連しています。胃の運動性と腸液の分泌も同様です。 5-HT 3 および 5-HT 4 受容体も胃腸の運動性に関連しているようです。
セロトニンは心血管系、平滑筋に作用し、血小板凝集を促進する血管作動性アミンであり、CNS で神経伝達物質として作用することは言うまでもなく、特に 大脳辺縁系 に関連し、不安、恐怖、うつ病などの反応を制御します。 、睡眠と痛みの知覚。
さまざまな行動状態では、この神経伝達物質のレベルに細胞外の変化が起こります。セロトニンレベルが低下すると、痛み、探索行動、運動活動、攻撃的および性的行動に対する感受性が高まることが知られています。男性と動物の両方において、精神障害は、注意欠陥に加えて、攻撃的で強迫的な行動などのセロトニン機能の変化と相関しています。
性行動に関して言えば、この神経伝達物質は視床下部からのゴナドトロピンの放出を阻害する役割を果たし、その結果性的反応が低下します。しかし、セロトニン作動性の低下により、性行為が容易になります。

セロトニンは、CNS 内の満腹中枢が位置する視床下部腹内側核の主な阻害剤です。この視床下部への影響は炭水化物に非常に特異的であり、タンパク質や脂質に作用するには他の補因子が必要です。したがって、セロトニンが減少すると体重が増加します。逆に高くなると食欲不振を引き起こします。
現在、セロトニンのレベルを上昇させる広く使用されている薬剤はフルオキセチンで、これはうつ病の治療に加えて、空腹感のコントロールを強化します。
獣医学に応用された薬理学 – ヘレニス・デ・ソウザ・スピノサ、シルヴァナ・リマ・グルニアック、マリア・マルタ・ベルナルディ。第4版。編集者グアナバラ・クーガン、2006 年。

ギャラリー