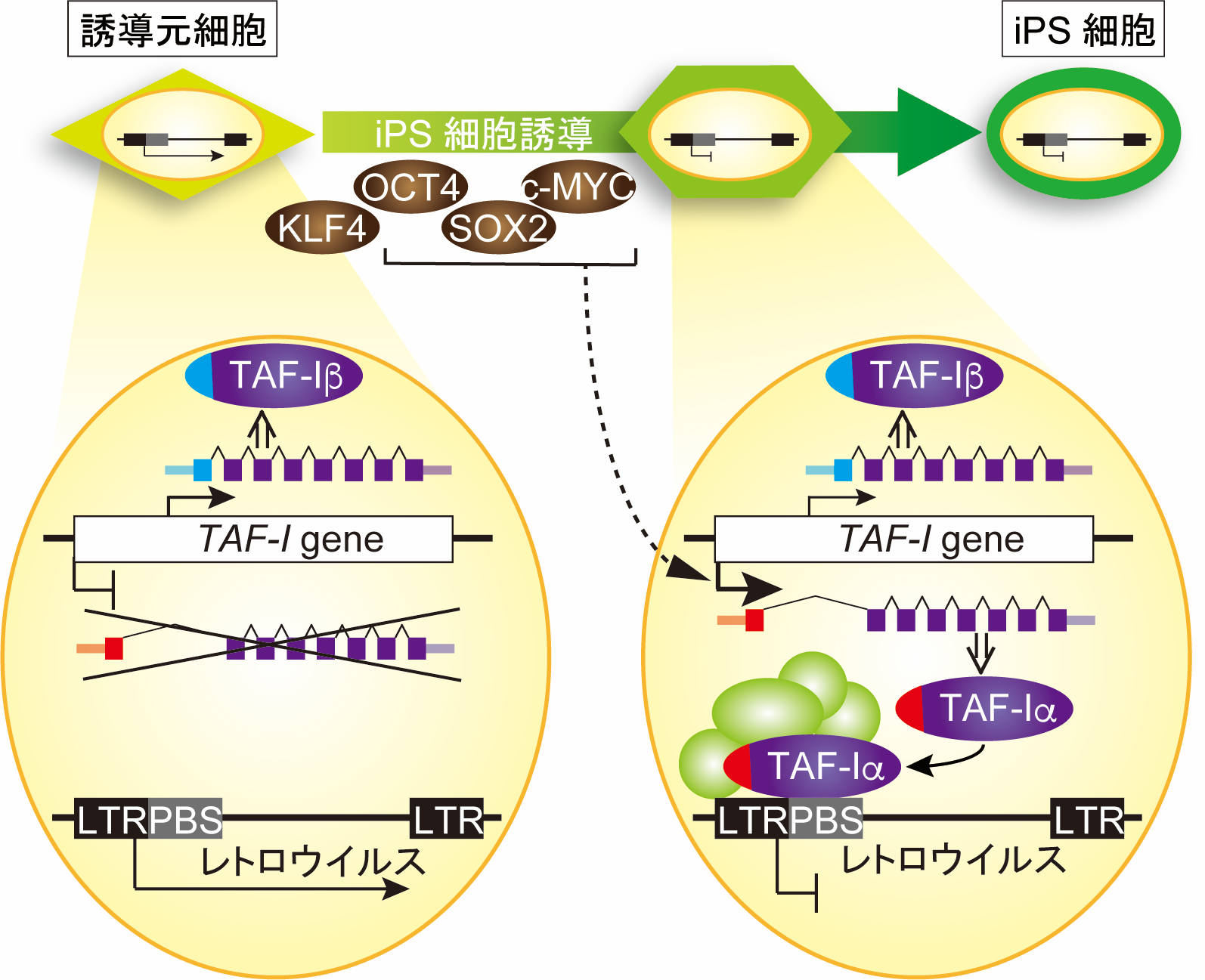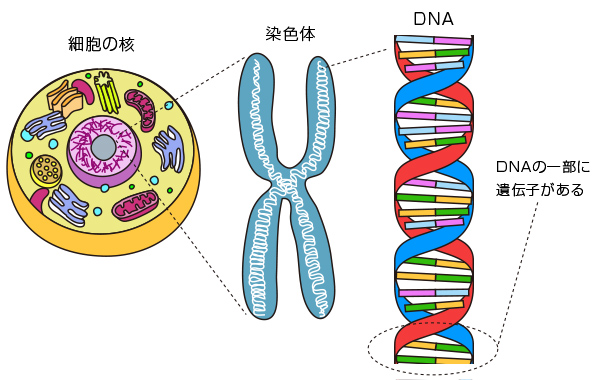
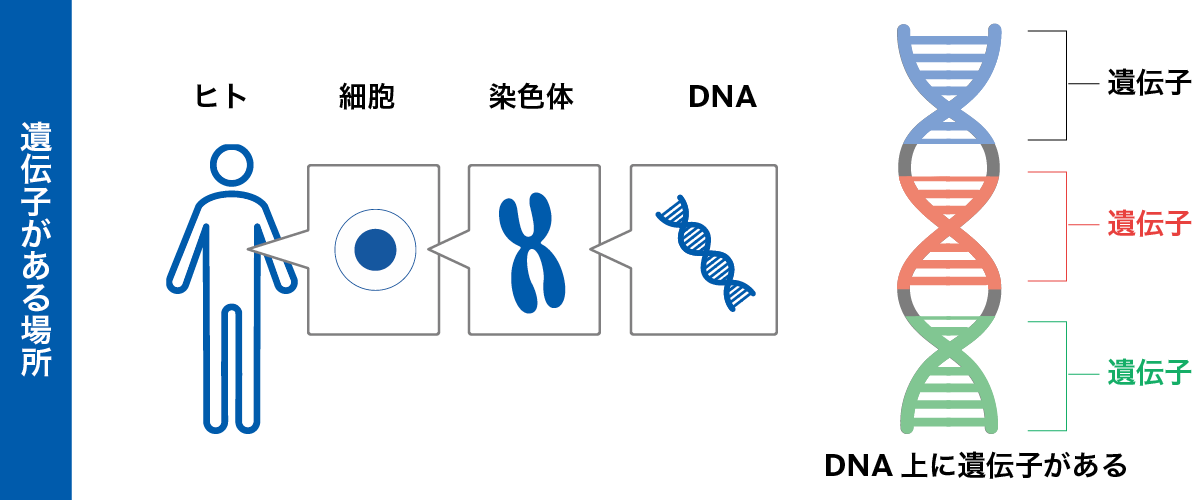
遺伝子は 、ポリペプチド鎖内の アミノ酸 生成のためのコードと発現のための調節配列を含む DNA 分子のセグメントですが、 ヒトゲノム には大きな非コード配列が存在します。コード配列は エクソン と呼ばれます。それらには、 イントロン と呼ばれる非コード領域が点在しており、最初は核内で RNA に転写されますが、 細胞質 内の最終 mRNA には存在せず、最終的なタンパク質産物にも現れません。多くの遺伝子では、エクソンの累積サイズはイントロンの累積サイズよりもはるかに小さくなります。

トンプソンによれば、典型的な状況では、ポリペプチドであれ機能的 RNA 分子であれ、機能的産物の生産に必要な染色体 DNA の配列として遺伝子を定義します。これは遺伝子の分子定義です。
すべての 真核生物 にはイントロンがありますが、 細菌 ではイントロンが稀であるか、存在しません。それらの存在の明らかな利点により、単一の遺伝子から複数のタンパク質産物を生成する代替処理が可能になります。
エクソンは短い傾向がありますが (約 150 ヌクレオチド )、イントロンは数百ヌクレオチドの長さになる場合があります。この不均衡により、高等真核生物ではエクソンよりもイントロンでの 組換え の頻度が高くなります。代替プロセスにより、元の遺伝子産物を破棄することなく、新しいエクソンの組み合わせをテストできます。
実際にコードする配列に加えて、遺伝子には、通常のメッセンジャー RNA 分子と同様に、遺伝子の正しい発現に必要な隣接するヌクレオチド配列も含まれており、その正しい量、正しい位置、 細胞周期 の正しい時間に存在します。
これらの隣接する配列は、遺伝子の 転写 の「開始」および「終了」シグナル配列を指します。 5′ 末端には転写が始まるプロモーター領域があり、3′ 末端には非翻訳領域である終結領域があります。
遺伝子とペプチドの関係
各遺伝子は異なるポリペプチドを指定し、それが生物の 表現型 に影響を与えるように作用します。ポリペプチドは、アミノ酸の直鎖状かつ特異的な鎖で構成される高分子です。ポリペプチドは タンパク質 の基本構成要素です。一部のタンパク質は生化学反応の 触媒 や 酵素 として機能します。他のものは細胞の構造要素を形成します。および他のものは、細胞の内外への物質の輸送を担当します。
遺伝子に 突然変異が 発生すると、そのポリペプチド産物が変化したり、生成されなくなったりして、生物内でのその役割が変化します。ホモ接合性の状態では、劣性突然変異は一般にポリペプチドの活性を低下させるか消失させます。
参考文献
遺伝学の基礎 / D. Peter Snustad、Michael J. Simmons。リオデジャネイロ:グアナバラ・クーガン、2008年。
遺伝子の分子生物学 / ジェームス D. ワトソン … [他]。ポルト アレグレ: アルメッド、2006
生物学 / ホセ・マリアーノ・アマビス、ジルベルト・ロドリゲス・マルソ。サンパウロ: モデルナ、2004
http://web.itu.edu.tr/sariel/publications.php
ギャラリー