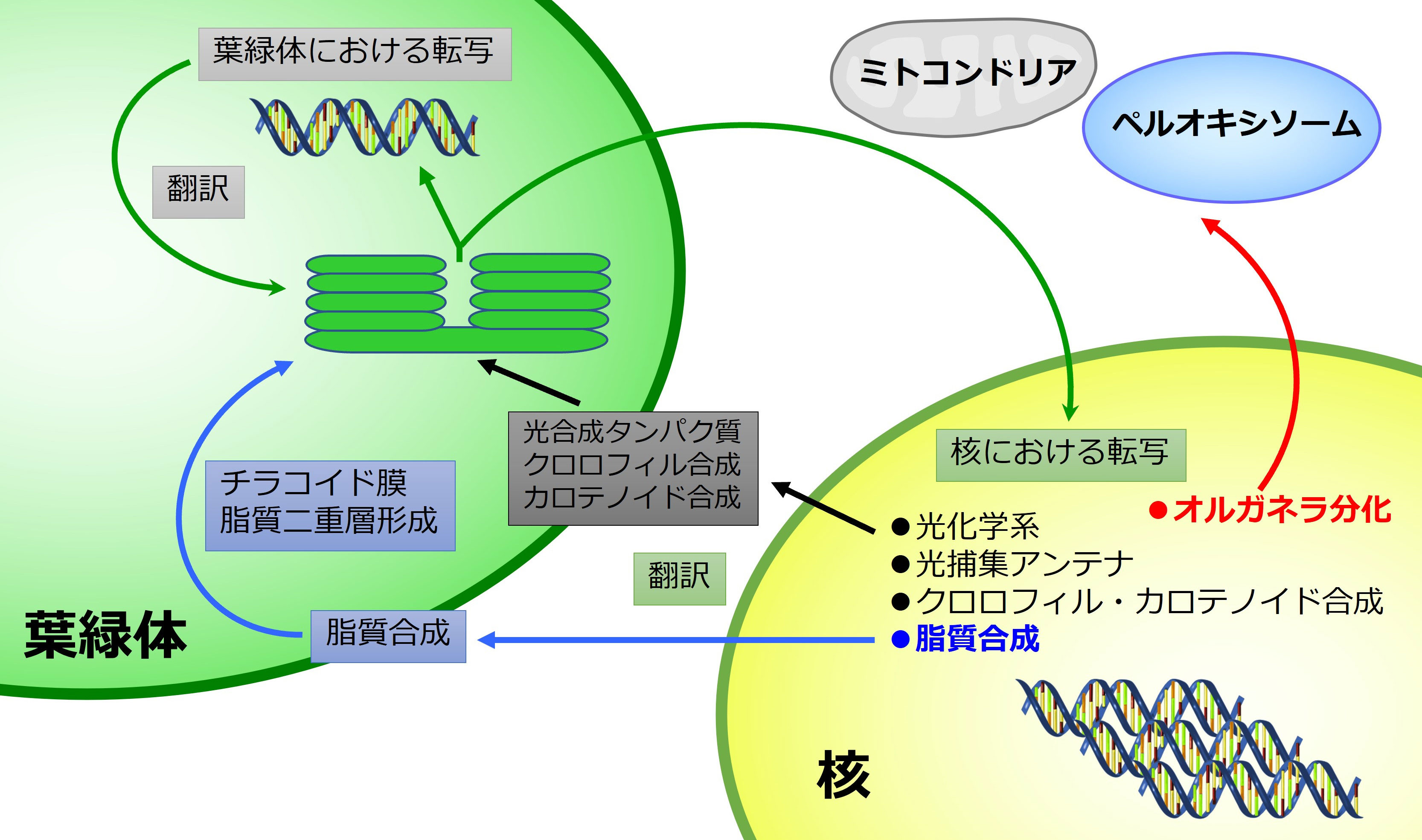
色素体は 、 原核 細胞 ( 藻類 など) および 真核細胞 (主に 植物細胞 ) の内部に存在する細胞小器官です。進化的に、これらの細胞小器官は 原核 生物であり、他の細胞によって取り込まれ、それらとの 共生関係 を発展させたと考えられています。この理論は、すべての色素体が独自の独立した遺伝物質、つまり DNA の二重環状鎖 (原核細胞で見られるものと同様) を持っているという事実によって補強されます。
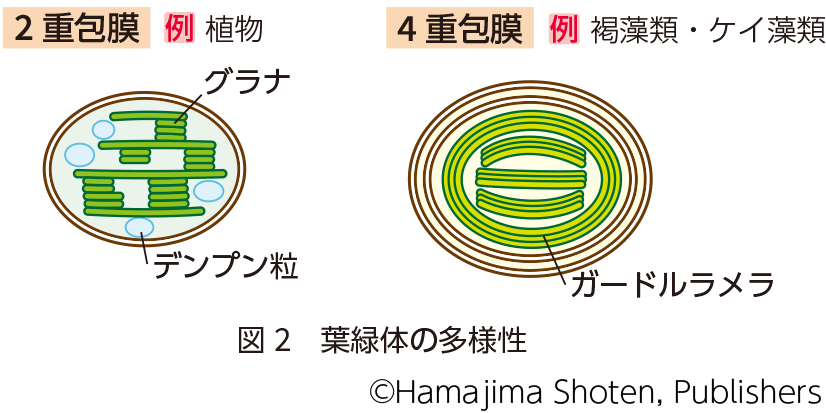
色素体の他の 細胞小器官 への分化は、それが存在する細胞の種類とその細胞の位置、色素体の化学組成、機能性および環境因子との相互作用などの一連の要因に依存します。植物細胞では、分裂組織領域に存在する色素体 (プロトプラスチドと呼ばれる) は、 葉緑体 、エティオププラスト、ジェロント プラスト、色素体 および 白質体 を形成できます。 葉緑体は 緑色の色素体であり、光にさらされると クロロフィル を合成し、 光合成 プロセスに不可欠です。エティオプラストは、まだ光にさらされていないため、基本的には活性色素を持たない 葉緑体 です。これらは通常、暗闇で成長する 被子植物 で発生し、黄色がかった色を引き起こしますが、植物が照らされると 葉緑体 に変換される可能性があります。それらは内部に幾何学的に組織化された構造、つまり クロロフィル の前駆体 分子 を含むプロラメラ体を持っています。ジェロントプラストは、植物の老化した部分に存在し、 アポトーシス (プログラムされた 細胞死 ) を開始する細胞内の光合成装置の分解に関連する特殊な細胞小器官です。色素体は、 カロテノイド などの クロロフィル 以外の色素の合成と貯蔵に特化しています。白血球は、予備物質の貯蔵と構造 有機化合物 の生成に関連する無色の色素体です。また、 アミロプラスト 、エライオプラスト、プロテノプラスト、タノソーマなどの他の形態に分化することもあります。これらの形態での分化は、ほとんどの場合、決定的ではなく、色素体の他の形態への再構築および再分化が発生する可能性があります。
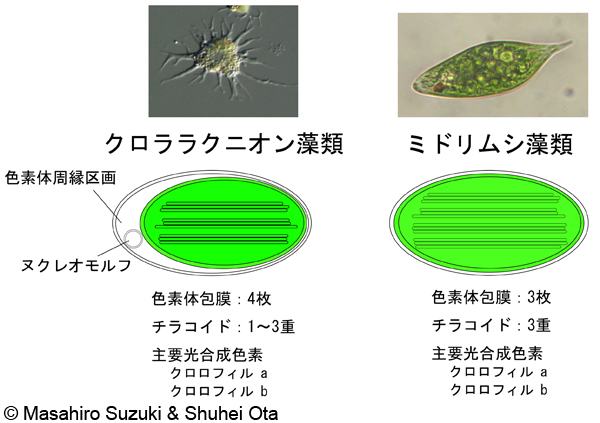
藻類 、原核生物、色素体には特定の特殊性があります。それらはすべてピレノイド分子を含み、色素体への分化はなく、白緑体は アミロプラスト への分化はありません。さらに、 藻類 の一部のグループは、色素体に由来する特定の形態を持っています。例としては、 赤色藻類 (紅藻類) のロードプラストが挙げられます。フィコシアニンは、光スペクトルの赤色帯域を使用して高い光合成効率を可能にする赤色色素であるフィコシアニンを合成して貯蔵します。また、ロードプラストは、樹木の枝に似た複雑なポリマー構造で知られる 樹上 デンプン と呼ばれる予備構造を合成し、細胞質に放出します。
色素体は、両親の一方の配偶子を介してのみ世代間で伝達されます(真核生物の 有性生殖 で起こる母親のミトコンドリアの遺伝と同様です)。 裸子植物では、 子孫は雄の 花粉 に存在する色素体を継承し、 被子植物 では、生存可能な子孫を形成する 2 つの 種 間で交配が起こる場合の双親継承の一部の例外を除いて、雌の配偶子を通じて継承が起こります。
参考文献:
宮村晋、黒岩哲、永田哲、1990 年。Triticum aestivum の前色素体から葉緑体とエチオプラストが発生する際の色素体核様体の増殖と分化。植物と細胞の生理学、31(5)、pp.597-602。

レイブン、PH、エバート、RF、アイヒホルン、SE、2005。植物の生物学。マクミラン。
Svab, Z.、Hajdukiewicz, P.、および Maliga, P.、1990。高等植物における色素体の安定した形質転換。米国科学アカデミー紀要、87(21)、8526-8530ページ。

Wolfe, KH、Morden, CW および Palmer, JD、1992 年。非光合成寄生植物からの最小色素体ゲノムの機能と進化。米国科学アカデミー紀要、89(22)、10648-10652ページ。
ギャラリー